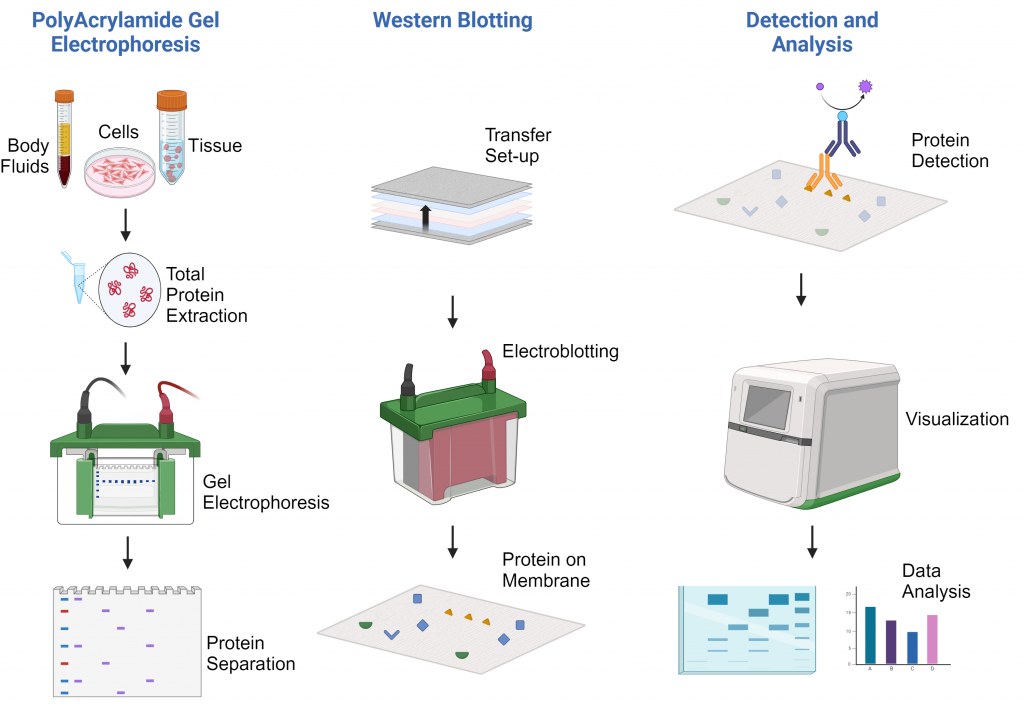
Western Blot-protocol

Principe van de techniek
Het principe van Western blotting omvat verschillende cruciale stappen:
-
Eiwitseparatie: Eiwitten worden op basis van grootte gescheiden door middel van SDS-PAGE.
-
Overdracht: Gescheiden eiwitten worden overgebracht naar een vast membraan (nitrocellulose of PVDF).
-
Blokkering: Membraanoppervlakken worden geblokkeerd om niet-specifieke binding te voorkomen.
-
Detectie: Specifieke primaire antilichamen herkennen doeleiwitten, waarna gelabelde secundaire antilichamen worden toegevoegd.
-
Visualisatie: Eiwit-antilichaamcomplexen worden zichtbaar gemaakt door middel van chemiluminescente, fluorescerende of colorimetrische reacties..
Het succes van deze techniek hangt in hoge mate af van de specificiteit van de antilichamen en de integriteit van de monstervoorbereiding.
De eerste belangrijke stap die niet over het hoofd mag worden gezien bij western blotting is de voorbereiding van het monster. Om reproduceerbare resultaten te verkrijgen, moet een efficiënte eiwitextractie en -zuivering worden uitgevoerd met behulp van een geschikte homogenisatiemethode die de intracellulaire inhoud van de cel efficiënt kan vrijmaken door de celmembranen te breken.
Eiwitisolatie uit verschillende soorten monsters
Eiwit extractie en isolatie zijn cruciale eerste stappen in een breed scala aan biochemische en moleculaire biologische technieken, waaronder Western blotting, massaspectrometrie, enzymatische testen en immunoprecipitatie. Het succes van downstreamtoepassingen hangt in hoge mate af van de kwaliteit, integriteit en representativiteit van de geïsoleerde eiwitmonsters.
Biologische monsters zoals gekweekte cellen, weefsels, bacteriën, gist en plantmateriaal vormen elk een unieke uitdaging voor eiwitextractie. Daarom is het kiezen van een geschikte isolatiestrategie essentieel om de opbrengst te maximaliseren, de functionaliteit van eiwitten te behouden en afbraak of modificatie tot een minimum te beperken..
1. Homogenisatie
-
Kies een homogenisatiemethode op basis van het type monster:
-
Ultrasoonbehandeling (voor delicate monsters)
-
Franse pers (voor bacteriën, gist)
-
Glasparelmaling (voor taaie cellen, schimmels)
-
Dounce/Potter-Elvehjem homogenisator (voor zachte weefsels)
-
Handmatig malen met vijzel en stamper (voor bevroren weefsels)
-
-
Doel: Efficiënte celyse zonder eiwitafbraak.
2. Voorbeeld van behandeling na afhandeling
-
Na blootstelling aan xenobiotica of genetische manipulatie (bijv. siRNA-knockdown):
-
Vries monsters onmiddellijk in vloeibare stikstof.
-
Lysaat onmiddellijk om endogene proteaseactiviteit te voorkomen..
-
-
Houd monsters tijdens de verwerking altijd koel..
3. Beheer van vries-/dooicyclus
-
Vermijd meerdere vries-/dooicycli om de integriteit van de eiwitten te beschermen.
-
Verdeel monsters in aliquots om onnodig ontdooien te voorkomen.
-
4. Selectie van lysisbuffer
a.Buffer afstemmen op locatie van doeleiwit:
-
Cytosolische eiwitten → Milde lysebuffer (bijv. NP-40, Triton X-100)
-
Membraangebonden eiwitten → Sterkere buffers (bijv. RIPA)
-
Nucleaire/mitochondriale eiwitten → Gespecialiseerde buffers (bijv. hypotonische buffer, Drosophila-homogenisatiebuffer)
b. Buffer afstemmen op gevoeligheid van antilichaam:
-
Detectie van natuurlijke eiwitten → Mild/ niet-ionisch detergent of geen detergent
-
Detectie van gedenatureerde eiwitten → Sterke/denaturerende omstandigheden
-
Voeg protease- en fosfatase-inhibitoren toe aan alle lysisbuffers.
c.Enkele veelgebruikte ingrediënten en waarom ze worden gebruikt:
-
HEPES: Houdt de pH-waarde stabiel.
-
Sucrose / Mannitol: Osmotische stabilisatoren om het barsten van organellen te voorkomen.
-
EDTA / EGTA: Chelaatvorming van tweewaardige kationen om metalloproteasen te remmen en afbraak te voorkomen.
-
Protease en fosfatase remmers: Beschermen eiwitten tegen enzymatische afbraak.
5. Kritische opmerkingen over monsterfractionering
a.Pas op voor eiwitverlies tijdens centrifugeren:
-
Test of het doeleiwit in het pellet (restanten) achterblijft..
-
Voorbeeld: Tot 50% van de myosine en ~66% van CSQ2 ging verloren in afval als dit niet werd gecontroleerd..
b.Controleer altijd of het eiwit van belang is:
-
Oplosbaar (cytoplasmatisch)
-
Onoplosbaar (cytoskelet)
-
Membraangebonden (organellaal)
Elektroforetische scheiding
Na eiwitextractie en kwantificering is de volgende cruciale fase in proteoomanalyse elektroforetische scheiding. Gelelektroforese blijft een fundamentele techniek voor het verbeteren van selectiviteit en gevoeligheid in proteoomonderzoek, waardoor complexe eiwitmengsels op basis van fysisch-chemische eigenschappen kunnen worden gescheiden.

Er zijn verschillende gelmatrices ontwikkeld voor elektroforetische toepassingen, waaronder agarose, zetmeel en polyacrylamide. Hiervan is polyacrylamide de matrix bij uitstek voor eiwitanalyse vanwege de mechanische stabiliteit, chemische inertie en het vermogen om de polymerisatie nauwkeurig te reguleren met bisacrylamide. Een veelgebruikte acrylamide:bisacrylamide-verhouding van 37,5:1 maakt het mogelijk om robuuste gels met reproduceerbare poriegroottes te genereren, die geschikt zijn voor eiwitfractionering met hoge resolutie.
Er worden twee hoofdvormen van polyacrylamide-gelelektroforese (PAGE) op grote schaal gebruikt: native PAGE en denaturerende SDS-PAGE.
-
Native PAGE behoudt de tertiaire en quaternaire structuren van eiwitten, waardoor ze kunnen worden gescheiden op basis van een combinatie van netto lading, hydrodynamische grootte en conformatie. In alkalische buffersystemen krijgen de meeste eiwitten een netto negatieve lading, waardoor ze zonder denaturatie naar de anode kunnen migreren. Een groot voordeel van native PAGE is dat de enzymatische activiteit en de natuurlijke eiwit-eiwitinteracties tijdens de scheiding behouden blijven.
-
SDS-PAGE (natriumdodecylsulfaat-polyacrylamide-gelelektroforese) is de meest gebruikte methode voor het scheiden van gedenatureerde eiwitten uitsluitend op basis van molecuulgewicht. Het anionische detergens SDS bindt zich gelijkmatig langs de polypeptideketen, waardoor een consistente negatieve lading ontstaat die evenredig is aan de lengte van het eiwit. Onder denaturerende omstandigheden (meestal door verhitting in 0,1% SDS) worden eiwitten gelineariseerd en nemen ze een langwerpige staafvorm aan, waardoor ze op basis van hun grootte door de polyacrylamidematrix kunnen migreren. Kleinere eiwitten passeren de gel sneller dan grotere eiwitten. Bovendien profiteert SDS-PAGE van isotachoforese, waardoor eiwitmonsters worden geconcentreerd in strakke, gedefinieerde banden, wat de resolutie verbetert (in gecontroleerde laboratoriumomstandigheden wordt een concentratie van tien keer waargenomen).
Aandacht voor de voorbereiding van monsters, de samenstelling van buffers en de eigenschappen van gels is essentieel voor een optimale scheiding en reproduceerbaarheid. De componenten van typische monsterbuffers en hun respectieve functies zijn samengevat in tabel 1.
Tabel 1. Veelgebruikte buffers voor monstervoorbereiding. Locatie van het eiwit Buffer Doel
| Locatie van eiwitten | Buffer | Doel van de componenten (typische concentraties tussen haakjes) |
| Hele cel |
Niet-denaturerende, niet-ionische buffer: NP-40 buffer: 50 mM Tris pH 8,0, 150 mM NaCl, 1% NP-40 of Triton X-100 |
Tris-HCl (10–50 mM): buffering van oplossing (helpt de stabiliteit van eiwitten te behouden) NaCl (50–150 mM): vermindert niet-specifieke binding, handhaaft de ionsterkte NP-40/Triton X-100 (0,1–1%): niet-ionische detergentia, maakt cytosolische en membraaneiwitten oplosbaar zonder ze te denatureren, maakt membranen permeabel en verhoogt de oplosbaarheid van eiwitten, waardoor aggregatie wordt voorkomen |
|
Denaturerende buffer: RIPA-buffer: 150 mM NaCl, 1% NP-40 of Triton X-100, 0,5% natrium deoxycholaat, 0,1% SDS, 50 mM Tris, pH 8,0 |
Natriumdeoxycholaat (0,1–0,5%): ionisch detergens, lyseert cellen en maakt cellulaire en membraancomponenten oplosbaar SDS (0,1–1%): ionisch detergens, veroorzaakt membraanverstoring en lineariseert eiwitten door zich eraan te binden |
|
| Cytoplasmatisch (oplosbaar) |
20 mM Tris-HCl, pH 7,5 of NP-40-buffer | Zie hierboven |
| Cytoplasmatisch (gebonden aan het cytoskelet) |
PIPES-Triton-buffer: 10 mM PIPES, 50 mM KCl, 1% Triton X-100, 10 mM EGTA, 3 mM MgCl2, 2 M glycerol |
LEIDINGEN (10 M): buffering van oplossing KCl (50–150 mM): vermindert niet-specifieke binding, handhaaft ionsterkte EGTA (1–3 mM): remming van Ca2+ metalloproteasen MgCl2 (1–5 mM): vermindert niet-specifieke binding, heeft samen met KCl een stabiliserend effect op sommige complexen Glycerol (1–10%): stabiliseert eiwitten |
| Membraangebonden | RIPA-buffer | Zie hierboven |
| Mitochondriën | RIPA buffer | Zie hierboven |
| Kernenergie | RIPA buffer | Zie hierboven; opmerking: NP-40 en Triton X-100 kunnen geen kernmembranen lyseren. |
| Locatie van eiwitten | Buffer |
Het benodigde gelpercentage is afhankelijk van de grootte van het eiwit dat u wilt onderzoeken:
| Eiwitgrootte | Gelpercentage |
| 4–40 kDa | 20% |
| 12–45 kDa | 15% |
| 10–70 kDa | 12% |
| 15–100 kDa | 10% |
| 25–100 kDa | 8% |
| 60-210 kDa | 5% |
Gradiëntgels kunnen ook worden gebruikt.
| Eiwitgrootte | Gelpercentage |
| 5-200 kDa | 4-12 gradient |
| 4-200 kDa | 4-20 gradient |
| 3.5-110 kDa | 10-20 gradient |
Continue versus discontinue buffersystemen
Elektroforetische scheidingssystemen kunnen grofweg worden ingedeeld in continue en discontinue (ook wel multifasische of isotachoforetische) buffersystemen, die elk worden gekenmerkt door verschillende bufferconfiguraties en scheidingsdynamica.
1. Continue buffersystemen:
-
Dezelfde bufferionen en pH in het monster, de gel en de reservoirs.
-
Gel gemaakt van een enkele acrylamideconcentratie.
-
Eiwitten scheiden direct in de gel zonder te stapelen.
-
Belangrijk punt: De resolutie hangt sterk af van kleine, geconcentreerde monsterhoeveelheden.
2. Discontinue (multifasische) buffersystemen:
-
Twee verschillende buffers met verschillende pH-waarden en ionsterkte:
-
Stapelgel: Stapelgel: pH ~6,8, laag Tris–HCl (0,125 M), grote poriën.
-
Resolving gel: pH ~8,8, hoog Tris–HCl (0,375 M), kleine poriën.
-
-
Loopbuffer (bijv. tris-glycine) aangebracht boven de stapelgel.
3. Scheidingsmechanisme:
-
Cl⁻-ionen fungeren als leidende ionen; glycinaat als achterblijvende ionen.
-
Eiwitten concentreren zich in strakke banden in de stapelgel (“stapeleffect”).
-
In de scheidingsgel worden eiwitten gescheiden op basis van hun molecuulgewicht, aangezien zij de belangrijkste ionen worden.
4. Voordelen van discontinue systemen:
-
Maakt het laden van grotere volumes verdunde monsters mogelijk.
-
Superieure resolutie dankzij stapeling en verschillende poriegroottes van de gel.
-
Effectievere scheiding op basis van molecuulgewicht.
Eiwitoverdracht
Na elektroforese worden eiwitten overgebracht van een polyacrylamidegel naar een membraan. Deze stap is cruciaal om eiwitten te immobiliseren voor verdere detectie en analyse (bijvoorbeeld western blotting).
1. Membranen gebruikt voor eiwitoverdracht
Er zijn verschillende membranen beschikbaar, elk met specifieke voordelen.:
-
Nitrocellulose is het meest gebruikte membraan. Het heeft een hoge eiwitbindende affiniteit, kan zowel eiwitten als glycoproteïnen immobiliseren en is compatibel met chemiluminescentie, chromogene en fluorescerende detectiemethoden. Het is echter broos omdat het is afgeleid van cellulose die is behandeld met salpeterzuur.
-
PVDF (polyvinylideendifluoride) biedt een betere mechanische sterkte dan nitrocellulose, waardoor het ideaal is voor strippen en opnieuw proberen of voor het sequencen van eiwitten. PVDF kan echter soms een hogere achtergrondkleuring veroorzaken, waardoor extra optimalisatie nodig is.
2. Methoden voor eiwitoverdracht
Eiwitten kunnen met behulp van verschillende methoden van de gel naar het membraan worden overgebracht:
-
Capillaire overdracht is gebaseerd op passieve capillaire werking.
-
Diffusieoverdracht zorgt ervoor dat eiwitten langzaam naar het membraan kunnen diffunderen.
-
Elektroblotting is de snelste en meest complete methode voor overdracht.
Van deze methoden is elektroblotting de voorkeursmethode vanwege de snelheid en efficiëntie ervan.
Soorten elektroblotting
Er zijn drie soorten elektroblotting op basis van de methode van buffergebruik:
-
Droge elektroblotting maakt gebruik van droge componenten zonder toegevoegde vloeistoffen.
-
Semi-droge elektroblotting maakt gebruik van met transferbuffer doordrenkte filterpapiertjes tussen het membraan en de gel.
-
Nat elektroblotting dompelt de gehele gel-membraan sandwich onder in een transferbuffer, wat met name wordt aanbevolen voor het overbrengen van grote eiwitten..
1. Principes van elektroblotting
Tijdens elektroblotting:
-
Er wordt een elektrisch veld loodrecht op de gel aangebracht.
-
Eiwitten, die geladen zijn, migreren uit de gel naar het membraan.
-
De gel moet naar de negatieve elektrode (kathode) gericht zijn, terwijl het membraan naar de positieve elektrode (anode) gericht is.
-
Om een goede overdracht te garanderen, worden de gel en het membraan stevig op elkaar gedrukt, met filterpapier en vezelpads aan beide zijden om luchtbellen te voorkomen, die een ongelijkmatige overdracht kunnen veroorzaken.
a.Overdrachtsbuffers
Overdrachtsbuffers helpen de eiwitstructuur te behouden en verbeteren de beweging tijdens de overdracht. Veelgebruikte buffers zijn onder andere:
-
Towbin-buffer (25 mM Tris, 192 mM glycine, 20% methanol, pH 8,3) wordt veel gebruikt voor natte overdracht.
-
Bjerrum Schafer-Nielsen-buffer (48 mM Tris, 39 mM glycine, pH 9,2, 20% methanol) heeft de voorkeur voor semi-droge overdracht.
-
CAPS-buffer (60 mM Tris en 40 mM CAPS) wordt vooral gebruikt voor eiwitten met hoge iso-elektrische punten.
Methanol in de buffer helpt gelzwelling te voorkomen en verbetert de binding van eiwitten aan het membraan. In sommige gevallen kunnen methanolvrije buffers echter nog steeds sterke detectiesignalen produceren.
De rol van de componenten van de transferbuffer wordt beschreven in tabel 2.
Tabel 2. Samenstelling van de buffer voor het uitvoeren en overbrengen en de functie van elke component
| Potentiële component | Rol |
| Buffers in gebruik | |
| Acetate | Goede scheiding in het hogere massabereik (100–500 kDa) |
| Glycine | Zorg voor glycinaat-ionen om een achterkant te vormen tijdens elektroforese in stakinggel. |
| MES | Nodig om een relatief constante pH te handhaven. Levert MES-ionen om een staartvorming te creëren tijdens elektroforese. MES zorgt voor een grotere scheiding bij een lager massabereik (<50 kDa). MES SDS-buffer is sneller dan MOPS SDS-buffer vanwege de lagere pKa van MES in vergelijking met MOPS. |
| MOPS | Nodig om een relatief constante pH te handhaven. Levert MOPS-ionen om een staartvorming te creëren tijdens elektroforese. MOPS zorgt voor een betere scheiding in het midden- tot hogere massabereik. Het verschil in de beweging van MES- en MOPS-ionen beïnvloedt de stapeling, wat resulteert in een verschil in het eiwitscheidingsbereik tussen MES- en MOPS-buffers. |
| SDS | Helpt eiwitten in een netto negatieve lading te houden |
| Tricine | Nodig om een relatief constante pH te handhaven. Tricine vervangt glycine in de loopbuffer en levert tricine-ionen om tijdens elektroforese een achterkant te vormen. Tricine zorgt voor een efficiëntere stapeling en scheiding van eiwitten met een laag moleculair gewicht. Geschikt voor het scheiden van eiwitten met een laag moleculair gewicht (0,5-50 kDa). |
| Tris-HCl | Noodzakelijk om een relatief constante pH te handhaven. Levert chloride-ionen die tijdens elektroforese in stakinggel de voorste rand vormen. Waterstofionen geleiden elektriciteit. |
| Overdrachtsbuffers | |
| Tris-HCl | Noodzakelijk om een relatief constante pH te handhaven |
| Glycine | Bij afwezigheid van methanol helpt het voorkomen dat gel opzwelt. |
| Methanol | Voorkomt dat gel tijdens de overdracht opzwelt en verbetert de binding van eiwitten aan nitrocellulose. Verwijdert SDS uit eiwitten, waardoor deze beter aan nitrocellulosemembranen kunnen binden. Sommige laboratoria voegen geen methanol meer toe aan overdrachtsbuffers, omdat de resultaten zonder methanol niet significant verschilden van die met methanol. Sommige protocollen gebruiken 10% ethanol in plaats van 20% methanol. |
| SDS | SDS (tot 0,1%) in transferbuffer verhoogt de efficiëntie van de eiwitoverdracht, vooral voor grote eiwitten, maar kan de hoeveelheid eiwit die zich aan het membraan bindt verminderen. In nitrocellulosemembranen met grotere poriëngroottes (0,45 μm) kunnen kleine SDS-gedenatureerde eiwitten door het membraan gaan. |
| CAPS | Noodzakelijk om een relatief constante pH te handhaven. Aanbevolen voor de overdracht van eiwitten met een hoog moleculair gewicht (>150 kDa). |
| CAPS: (Cyclohexylamino)-L-propaansulfonzuur; MES: 2-(N-morfolino)ethaansulfonzuur; MOPS: 3-(N-morfolino)propaansulfonzuur; SDS: Natriumdodecylsulfaat; Tris-HCl: Tris-hydrochloride. |
|
b.Belangrijke overwegingen
-
Eiwitten met een zeer laag moleculair gewicht (<10 kDa) kunnen mogelijk niet efficiënt worden overgedragen..
-
Als de pH van de transferbuffer lager is dan het iso-elektrische punt van het eiwit, kunnen eiwitten terug migreren.
-
Voor het efficiënt overbrengen van eiwitten met een hoog iso-elektrisch punt zijn vaak speciale buffers (zoals CAPS-gebaseerde buffers) nodig.
Blokkerende stap bij Western blotting
Na een succesvolle eiwitoverdracht naar het membraan is blokkeren een cruciale stap om de specificiteit en duidelijkheid van de daaropvolgende immunodetectie te garanderen.

1.Doel en belang
-
Blokkering voorkomt niet-specifieke binding van antilichamen aan het membraan, waardoor achtergrondruis tot een minimum wordt beperkt en vals-positieve signalen worden geëlimineerd.
-
Het verbetert de nauwkeurigheid en betrouwbaarheid van eiwitdetectie tijdens western blotting.
2. Blokkeerprocedure
-
Het membraan wordt gedurende 1 uur bij kamertemperatuur geïncubeerd in een blokkerende oplossing die het volgende bevat:
-
Bovine Serum Albumine (BSA) of magere melkpoeder, verdund in:
-
Tris-gebufferde zoutoplossing met 0,1% Tween 20 (TBST), of
-
Fosfaatgebufferde zoutoplossing met 0,1% Tween 20 (PBST).
-
-
-
De eiwitten in BSA en melk (voornamelijk caseïne en wei-eiwitten) binden zich aan onbezette plaatsen op het membraan, waardoor niet-specifieke interacties tussen het membraan en de primaire of secundaire antilichamen worden voorkomen.
3. Membraaneigenschappen en rol van Tween 20
-
Nitrocellulose en PVDF-membranen vertonen een hoge affiniteit voor eiwitbinding, voornamelijk door hydrofobe interacties.
-
Hoewel BSA en melkeiwitten een zwakke binding kunnen aangaan met overgedragen eiwitten, vermindert de toevoeging van Tween 20 in de blokkerende buffer deze niet-specifieke interacties, waardoor een effectievere blokkering wordt gegarandeerd.
Incubatie en detectie van antilichamen in Western blotting
1. De rol van antilichaamincubering bij Western blotting
Na het blokkeren van het membraan om niet-specifieke binding te voorkomen, gaat western blotting over naar de cruciale stap van antilichaamincubatie. Eerst wordt het membraan geïncubeerd met een primair antilichaam dat zich specifiek bindt aan het doeleiwit. Nadat ongebonden antilichamen zijn weggewassen met buffers zoals TBST of PBST, wordt een secundair antilichaam toegevoegd. Dit secundaire antilichaam herkent het primaire antilichaam en is doorgaans gekoppeld aan een enzym zoals HRP (Horseradish Peroxidase) of Alkaline Phosphatase (AP), waardoor vervolgens signalen kunnen worden gedetecteerd.
-
Primaire antilichamen: Essentieel voor specifieke en selectieve binding aan doeleiwitten.
-
Secundaire antilichamen: maken detectie en versterking van het signaal mogelijk.
Tip: Het is belangrijk om de antilichaamconcentratie te optimaliseren om de gevoeligheid te maximaliseren en de achtergrond te verminderen. Onze technische ondersteuning kan u helpen met het optimaliseren van het protocol..
Wasstappen zijn cruciaal om ongebonden antilichamen te verwijderen. Overmatig wassen (>20 minuten) kan echter het signaal verzwakken — een evenwichtige timing is dus essentieel!
2. Signaaldetectiemethoden
Zodra het doelwitproteïne is gebonden door gelabelde antilichamen, moet het signaal worden gedetecteerd. Hiervoor zijn verschillende methoden beschikbaar:
a. Chemiluminescentie (ECL)
Verbeterde chemiluminescentie (ECL) is de meest populaire detectiemethode. Met behulp van HRP-geconjugeerde secundaire antilichamen en ECL-substraten wordt licht gegenereerd en vastgelegd op film of met een digitale beeldvormer.
-
Zeer gevoelig
-
Breed dynamisch bereik
-
Ideaal voor het detecteren van eiwitten in lage concentraties
Aanbevolen producten:
HRP-geconjugeerde secundaire antilichamen
ECL-substraatkits
ECL-detectiekits
b. Fluorescentiedetectie
Fluorescerende kleurstof-geconjugeerde secundaire antilichamen maken multiplexdetectie mogelijk, wat betekent dat u meerdere eiwitten tegelijk kunt visualiseren met behulp van verschillende fluorochromen..
-
Geschikt voor gelijktijdige detectie van meerdere doelen
-
Uitstekend dynamisch bereik en kwantificering
-
Het beste bij gebruik van fluorochromen in het infraroodgebied
Aanbevolen producten:
Secundaire antilichamen (zichtbaar en infrarood bereik)
Fluorescentiebeeldvormingssystemen
c. Chromogene detectie
Chromogene methoden maken gebruik van enzymreacties om een gekleurd neerslag op het membraan te genereren, dat met het blote oog zichtbaar is.
- Ideaal wanneer de beeldvormingsapparatuur beperkt is.
-
Eenvoudige visualisatie
-
Geen speciale apparatuur nodig
-
Ideaal voor educatieve laboratoria en fundamenteel onderzoek
Aanbevolen producten:
Chromogene substraatkits (e.g., BCIP/NBT)
d. Radioactieve detectie (tegenwoordig minder gebruikelijk)
Radioactieve sondes werden vroeger veel gebruikt voor western blotting, maar zijn nu grotendeels vervangen door veiligere, even gevoelige methoden zoals ECL en fluorescentie. Het kiezen van de juiste detectiemethode is cruciaal voor het succes van uw western blot en hangt af van de gevoeligheid, specificiteit en het type analyse dat u nodig hebt:
-
Maximale gevoeligheid? → Kies Verbeterde chemiluminescentie (ECL) detectie.
ECL biedt hoge gevoeligheid en een breed dynamisch bereik, waardoor het ideaal is voor het detecteren van eiwitten in lage concentraties. De signaalintensiteit kan worden vastgelegd op röntgenfilms of digitale beeldvormingssystemen.. -
Multiplexanalyse (simultane detectie van meerdere doelen)? → Kies fluorescentiedetectie.
Fluorescerende secundaire antilichamen maken gelijktijdige detectie van meerdere eiwitten mogelijk zonder dat membranen moeten worden gestript en opnieuw moeten worden onderzocht. Signalen zijn zeer stabiel, kwantitatief en ideaal voor vergelijkende studies.. -
Eenvoudige, snelle visualisatie? → Kies voor chromogene detectie.
Chromogene systemen maken gebruik van enzym-substraat reacties om een gekleurd neerslag direct op het membraan te produceren. Hoewel ze minder gevoelig zijn dan ECL of fluorescentie, zijn ze gemakkelijk te gebruiken en vereisen ze geen gespecialiseerde beeldvormingsapparatuur..
Bovendien moeten de keuze van primaire en secundaire antilichamen, evenals substraten of detectiereagentia, compatibel zijn met de door u gekozen detectiemethode. Een juiste combinatie is essentieel om specifieke en sterke signalen te verkrijgen en achtergrondruis tot een minimum te beperken..